
In an article yesterday, I gave a short overview of the arrangement and structure of muscles. Here, I will describe the biochemistry of muscle contraction. Readers may find it slightly easier to follow the discussion that follows by first viewing this short animation, which describes the sliding filament model of muscle contraction.
The Structure of a Muscle Fiber
I previously noted that muscles contain thousands of cylindrical cells called muscle fibers, or myocytes. The motor neuron terminates at the muscle fiber’s neuromuscular junction. The tip of the motor neuron is known as the axon terminal, and it contains sacs of acetylcholine, an important neurotransmitter involved in muscle contraction. The muscle fiber also has a membrane called the sarcolemma, containing acetylcholine receptor sites, in addition to an inactivator called cholinesterase. The small space between the sarcolemma and axon terminal is called the synapse, or synaptic cleft.
The muscle fiber contains thousands of individual contracting units known as sarcomeres. These are organized end-to-end in cylinders known as myofibrils. In the center of the sarcomere are thick filaments comprised predominantly of the protein myosin, and thin filaments containing actin can be found at the ends, attached to the end boundaries of the sarcomere (known as the Z discs) by the protein titin. The structure of the muscle fiber is shown in the figure below:
Image credit: OpenStax, CC BY 4.0 https://creativecommons.org/licenses/by/4.0, via Wikimedia Commons.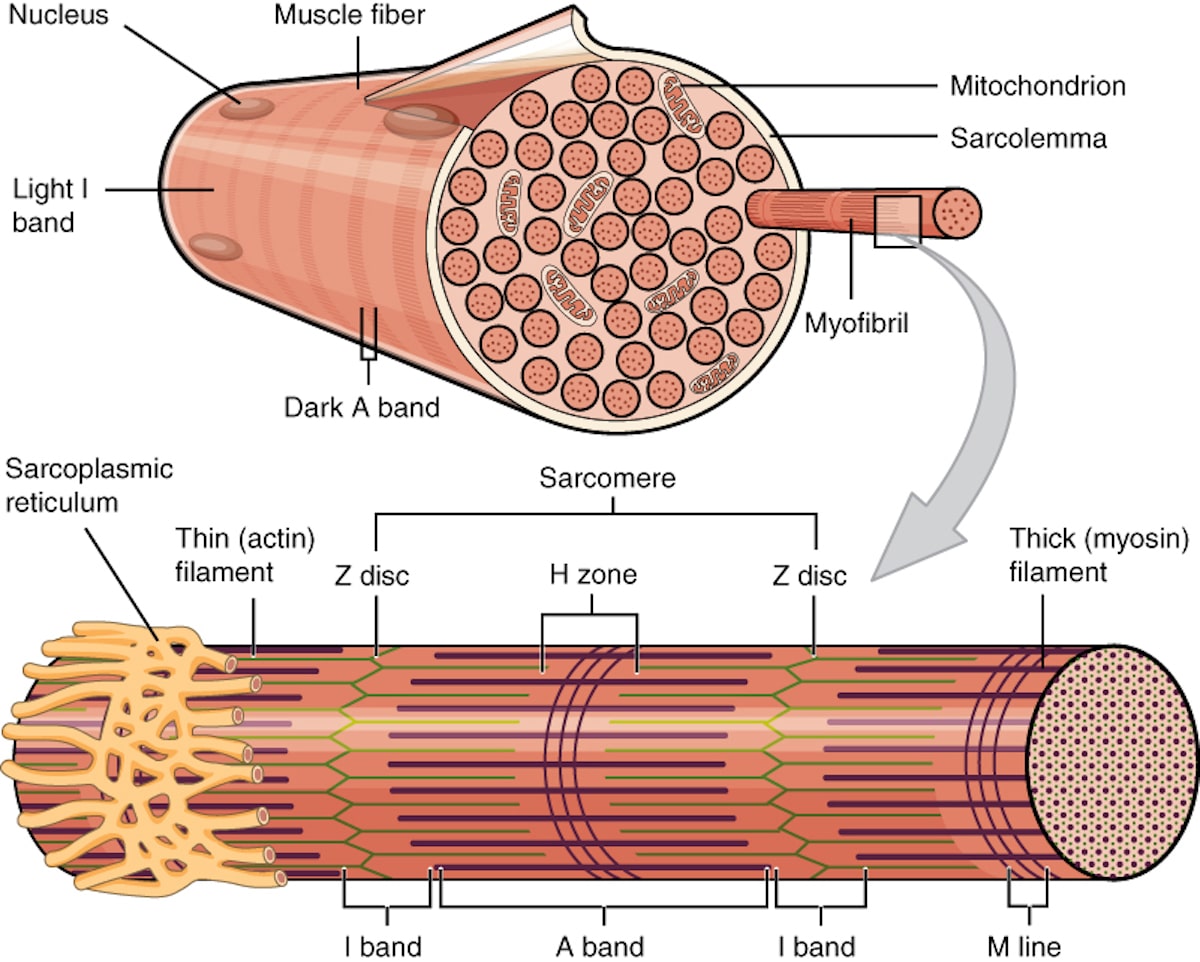
Image credit: OpenStax, CC BY 4.0 https://creativecommons.org/licenses/by/4.0, via Wikimedia Commons.
Muscle contraction is driven by two contractile proteins — myosin and actin. Each myosin molecule consists of a long tail and a globular head. Myosin heads have ATPase activity, which allows them to hydrolyze ATP to generate energy for muscle contraction. Myosin heads also have binding sites for actin and ATP. Actin has binding sites for myosin heads. However, these binding sites are typically covered by two inhibitory proteins known as tropomyosin and troponin when the muscle is relaxed. These inhibitory proteins prevent the sliding of myosin and actin during relaxation of the muscle fiber.
The sarcomeres are surrounded by the sarcoplasmic reticulum (the muscular equivalent of the endoplasmic reticulum), which serves as a reservoir of calcium ions (Ca2+). As we shall see, Ca2+ ions are required for muscle contraction.
Polarization of the Sarcolemma
When a muscle fiber is in a state of relaxation, the sarcolemma has a resting potential, or is said to be polarized. This refers to the difference in electrical charges between the inside and outside. When the sarcolemma is polarized, there is a positive charge outside relative to the negatively charged inside. There is a greater concentration of sodium ions (Na+) outside the cell and a greater concentration of potassium ions (K+) and negative ions inside the cell.
Because of the concentration gradient, the Na+ ions tend to diffuse into the cell and the K+ ions tend to diffuse outside. These are actively transported back out and in respectively by the sodium and potassium pumps, which depend upon ATP to maintain polarization and muscle relaxation until a change is stimulated by a nerve impulse.
Depolarization of the Sarcolemma
The first step in muscle contraction is the arrival of a nerve impulse at the axon terminal, stimulating the release of the neurotransmitter acetylcholine. The acetylcholine diffuses across the synapse and binds to acetylcholine receptors on the sarcolemma. This renders the sarcolemma extremely permeable to Na+ ions, which rapidly enter the cell. This reverses the charges such that there is now a positive charge on the inside of the sarcolemma relative to the outside. This charge reversal is known as depolarization.
Inward folds on the sarcolemma known as transverse tubules (or, T tubules) carry this electrical impulse (referred to as an action potential) to the interior of the muscle cell. Depolarization triggers the release of Ca2+ ions from the sarcoplasmic reticulum. These bind to the troponin-tropomyosin complex, moving it away from the actin filaments.
The Sliding Filament
With the binding sites on actin now available, actin can be bound by the myosin heads, forming cross-bridges. Once the cross-bridges are formed, the myosin heads pivot, pulling the thin filaments towards the center of the sarcomere. This action is called the power stroke and is powered by the energy released when ATP is hydrolyzed. After the power stroke, the myosin heads require ATP to detach from actin. ATP is hydrolyzed into ADP and inorganic phosphate, which energizes the myosin head for the next cycle.
The cycle of cross-bridge formation, power stroke, ATP hydrolysis, and detachment repeats as long as calcium ions are present and ATP is available. This results in the shortening of the sarcomere and, collectively, the entire muscle fiber. This leads to muscle contraction. The force generated by many muscle fibers contracting in unison allows for body movement. The sliding filament model is graphically summarized in the figure below:
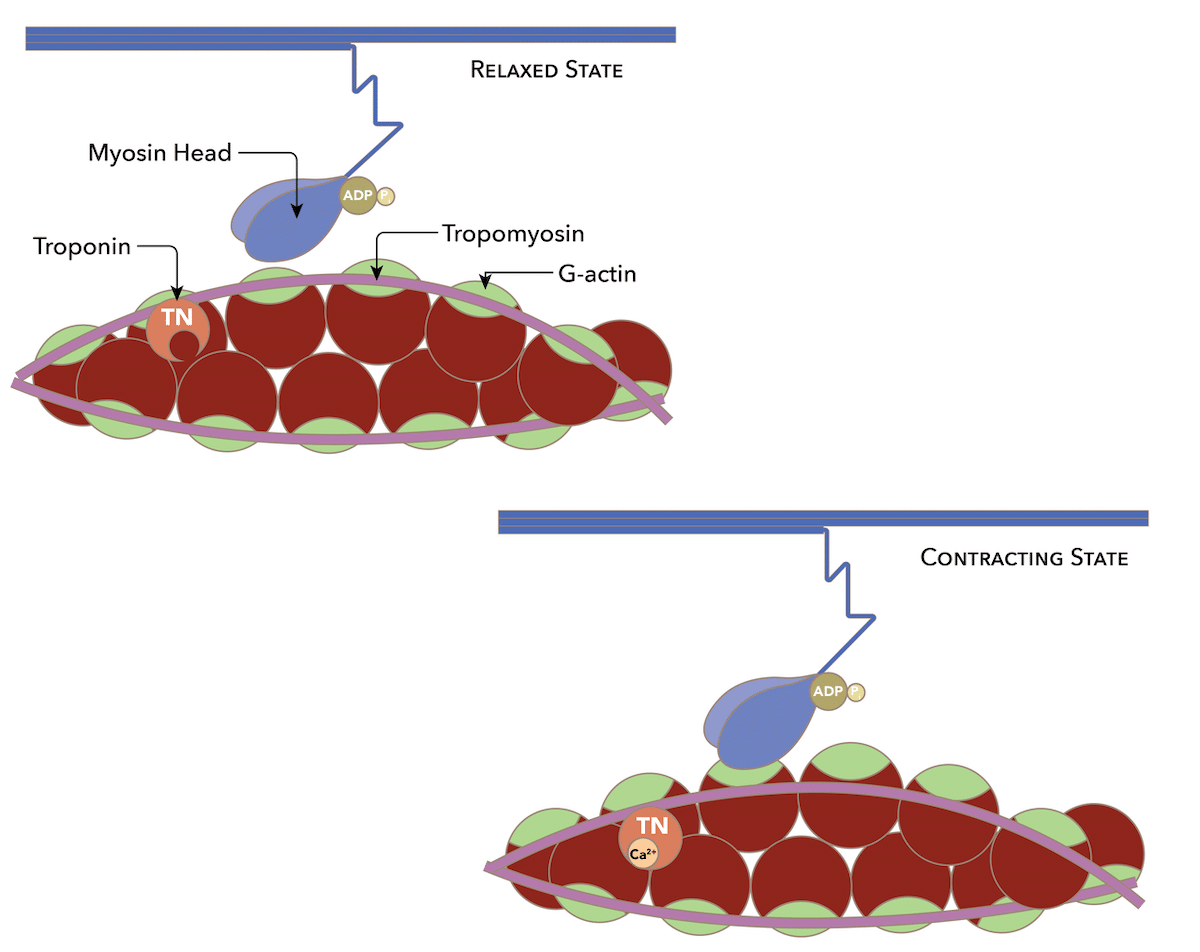
Image credit: Daniel Walsh and Alan Sved, CC BY-SA 4.0 https://creativecommons.org/licenses/by-sa/4.0, via Wikimedia Commons.
Repolarization
Muscle relaxation occurs when the electrical stimulation ceases — resulting in the ionic concentrations inside and outside the cell returning to their resting state. Potassium ions exit the cell, restoring the positive charge outside and negative charge inside the cell. Repolarization results in the myosin heads releasing their grip on actin, and calcium ions are actively transported back into the sarcoplasmic reticulum, causing muscle relaxation. Acetylcholine is inactivated by cholinesterase in the sarcolemma.
Following repolarization, sodium ions are pumped outside the cell and potassium ions are pumped inside. This re-establishes the negative resting membrane potential (polarization) and ensures that the sodium concentration outside the cell remains higher than inside, while the potassium concentration inside the cell remains higher than outside. This ensures that a state of electrochemical equilibrium remains unreached, and is crucial for maintaining the excitability of the cell and its ability to generate future action potentials.
Evidence of Design
As should be apparent from the forgoing discussion, muscle contraction — which we so easily take for granted — is an incredibly complex and elegant process, involving incredible engineering and design. The process of muscle contraction and relaxation requires the coordinated action of actin, myosin, troponin, tropomyosin, acetyl choline, ion channels, and much more. To contend that the phenomenon of muscle contraction arose through a blind and undirected process, one tiny Darwinian step after the other, seems to me to strain credulity.
Note: This article was originally published on October 27 2023 at Evolution News & Science Today.