I have previously published several articles in the past on the incredible design, and exquisite engineering, of the eukaryotic cell division cycle (see this recent article for links to previous essays on this subject). I also recently published a paper, in the journal BIO-Complexity, in which I documented significant obstacles to the origins of the eukaryotic cell cycle by evolutionary processes (available for free here). [1] Here, I will describe several aspects of the cell cycle that render it irreducibly complex, which are also discussed in my paper.
Condensins
Condensins are protein complexes that play a crucial role in the organization and segregation of chromosomes during cell division. They are highly conserved across eukaryotes. Condensin I is active during late prophase and contributes to the structural integrity of chromosomes following the break-down of the nuclear envelope. Condensin II functions earlier in prophase and is involved in the initial stages of chromosome condensation in the nucleus.
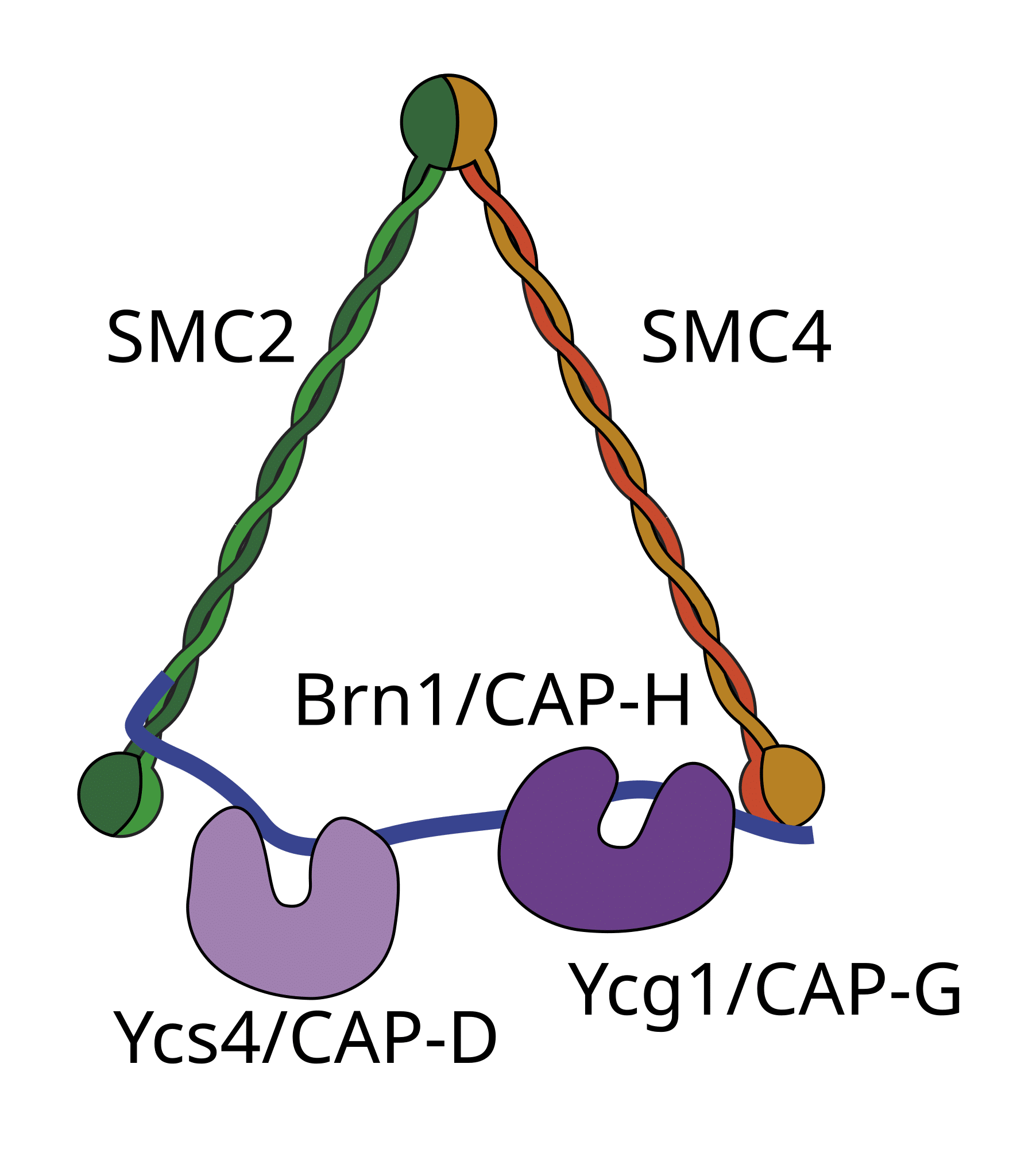
Image source: Wikimedia Commons.
Condensin molecules are composed of five subunits (as shown in the figure), including the SMC (Structural Maintenance of Chromosomes) proteins SMC2 and SMC4, which possess ATPase activity. SMC proteins possess coiled-coil domains (long, flexible arms that fold back on themselves, creating a V-shaped structure), a hinge domain that facilitates the dimerization of the two SMC proteins; and head domains containing ATP-binding and ATPase sites, energizing the activities of condensins. In addition to the SMC subunits, there are also three non-SMC subunits, which bind specific regions of DNA and assist in regulation of condensin activity.
Condensin complexes load onto chromatin in a stepwise manner, directed by non-SMC subunits. The SMC subunits create loops in DNA, utilizing their ATPase activity. These loops are stabilized and condensed into mitotic chromosomes.
The condensin proteins are crucial for the process of cell division. In their absence, the consequence would be chromosomal disorganization, as well as great difficulty in achieving proper segregation during mitosis.
Kinetochores
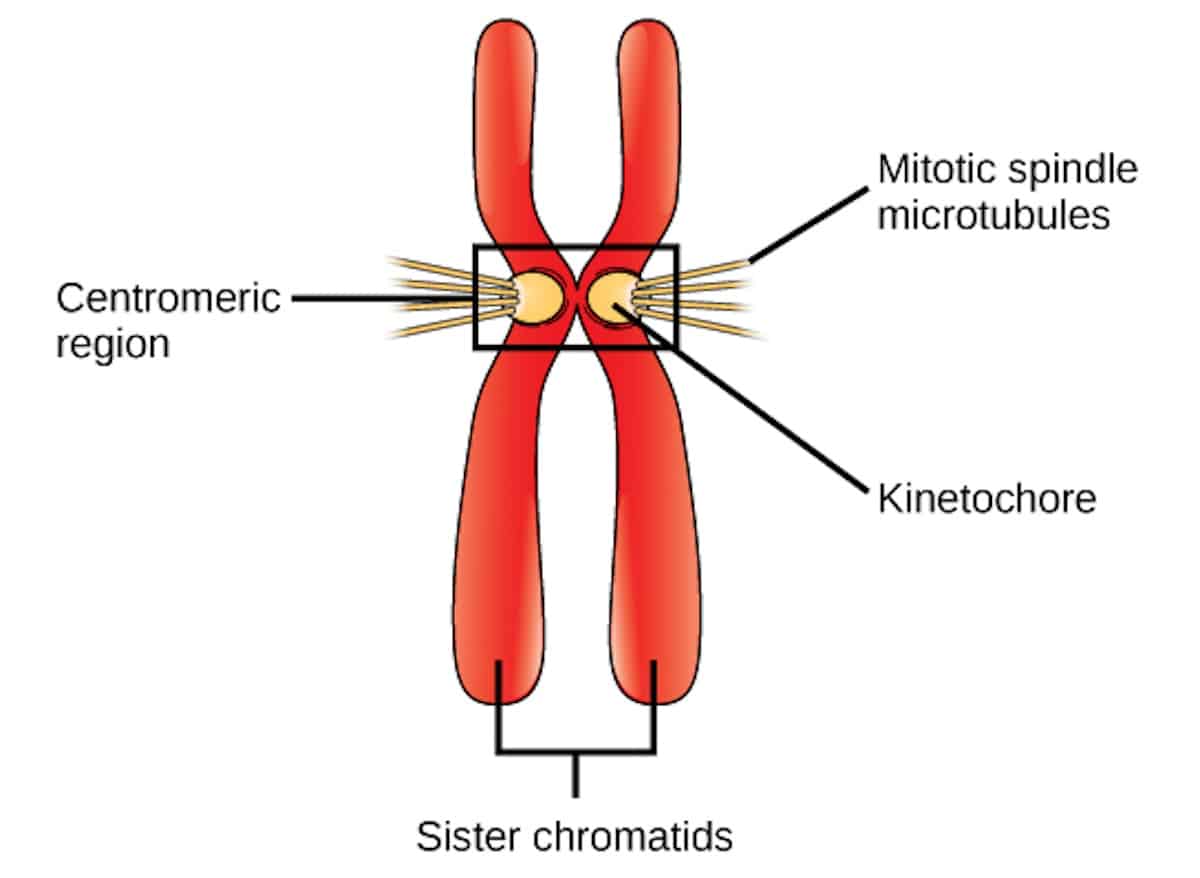
Image credit: CNX OpenStax, CC BY 4.0 https://creativecommons.org/licenses/by/4.0, via Wikimedia Commons.
A complex of proteins, known as the kinetochore, assembles around the centromere of each chromosome (as shown in the figure), and is critical to the process of mitotic cell division. Each kinetochore serves as an attachment site for the spindle microtubules, which radiate from the centrosomes at the cell’s poles. Kinetochores assist with the alignment of chromosomes at the equatorial plane of the cell during metaphase, ensuring equal distribution of genetic material. Kinetochores also sense tension generated by microtubule pulling, thereby ensuring proper attachment. If improper attachments occur (e.g. if the kinetochores of both sister chromatids are attached to the same pole), these errors can be corrected by the kinetochore-associated machinery.
What would be the consequence if there were no kinetochores? This would result in the improper attachment of the chromosomes to the spindle apparatus, and the genetic material would be unequally distributed to the daughter cells. Indeed, so critical are the kinetochores to the process of cell division that they are found ubiquitously throughout all known eukaryotic organisms.
Separase and the Anaphase Promoting Complex
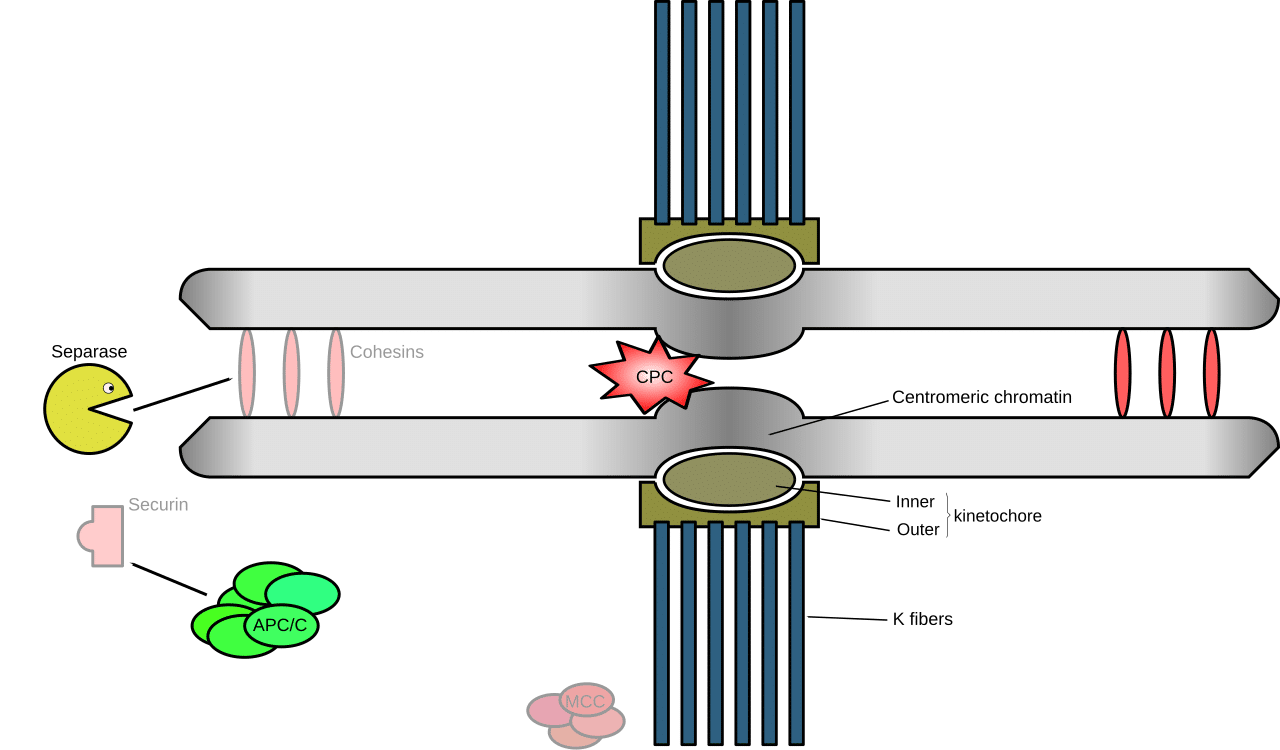
Image source: Wikimedia Commons.
Progression from metaphase to anaphase is mediated by the anaphase promoting complex or cyclosome (APC/C), an E3 ubiquitin ligase. When bound to its coactivator, Cdc20, the APC/C functions to ubiquitylate securin (a protein that prevents the cleavage of cohesin by the enzyme separase). Ubiquitylation of securin targets it for destruction by the cell’s molecular shredder, the proteasome. This liberates the enzyme separase to cleave the cohesin ring that tethers the sister chromatids together, thereby promoting sister chromatid separation.
In the absence of separase, the sister chromatids would fail to separate, and the cell would be rendered unable to segregate its chromosomes at anaphase. Indeed, experimental knockout studies have shown that deleting separase results in embryonic lethality. [2,3] Cell cycle progression would also be halted in the absence of the APC/C, inhibiting the progression from metaphase to anaphase. Indeed, experimental studies knocking out APC2 (a core APC/C subunit) in mice, for example, resulted in lethal bone marrow failure within only seven days. [4]
Aurora Kinases
Aurora kinases are also crucial to proper spindle formation and chromosome segregation. Aurora kinase A phosphorylates proteins involved in microtubule organization and facilitates the accurate attachment of microtubules to kinetochores. Indeed, “Aurora A null mice die early during embryonic development during the 16-cell stage. These Aurora A null embryos have defects in mitosis, particularly in spindle assembly, supporting critical functions of Aurora A during mitotic transitions.” [5] This indicates that Aurora kinase A is among the components that are essential for successful cell division.
Microtubules
I have previously described the critical role of microtubules in cell division. Microtubules radiate from centrosomes and anchor to the kinetochore complex, assembled around the centromere of each chromosome. During metaphase, the chromosomes are aligned along the equatorial plane of the cell, bound to microtubules at the kinetochore. In anaphase, the sister chromatids are pulled apart by the microtubules, driven by poleward spindle forces. The microtubules are, therefore, essential for segregating the sister chromatids into the two daughter cells.
In the absence of the microtubules, mitotic spindle assembly would thus be severely impaired, inhibiting chromosome alignment and segregation. Indeed, experimental studies with mouse embryos that are deficient in γ-tubulin exhibit a mitotic arrest that arrests development at the morula/blastocyst stages. [6]
The Contractile Ring
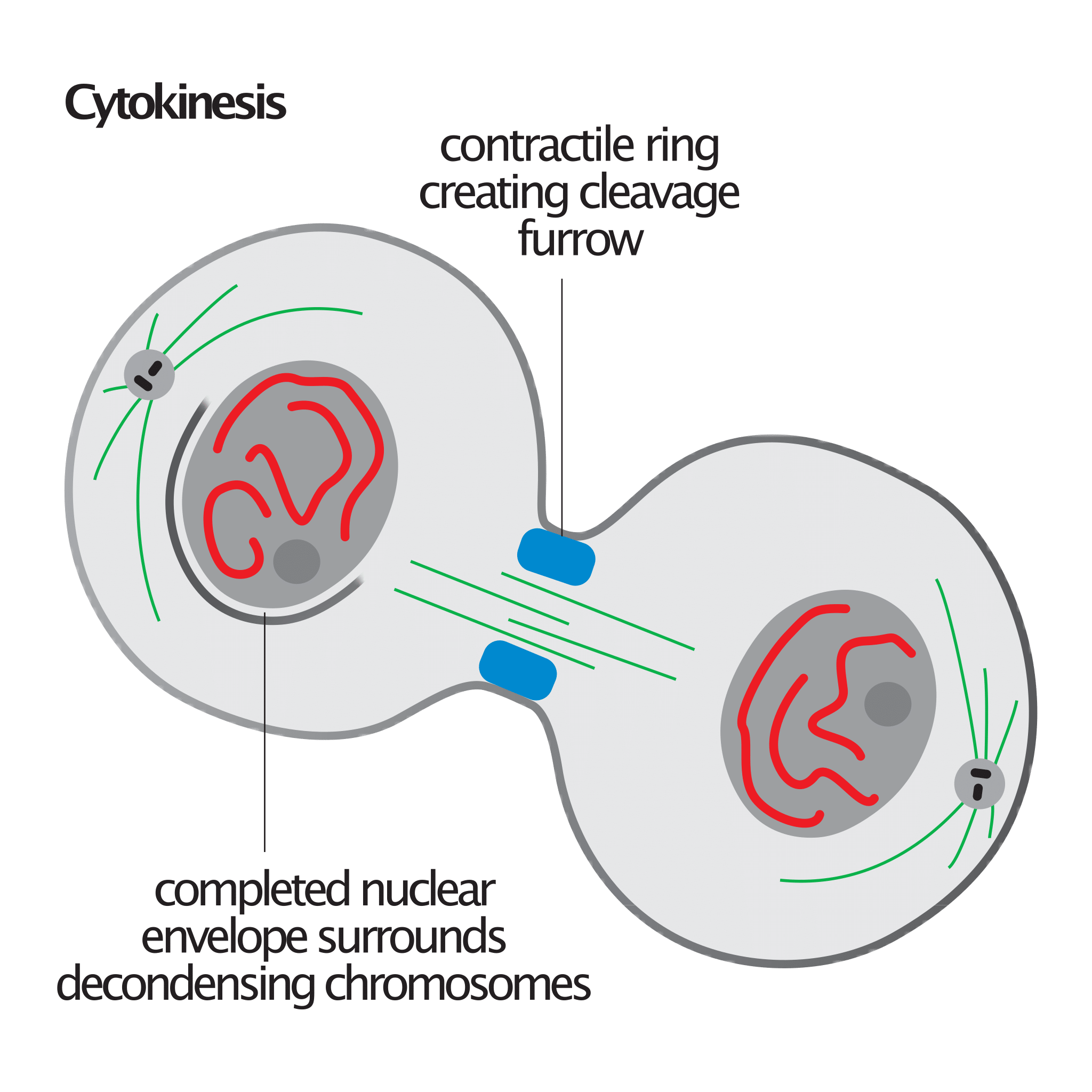
Image credit: David O Morgan, via Wikimedia Commons.
The contractile ring is also critical to the process of cytokinesis, the final stage of mitosis where the cell physically divides into two daughter cells. It is principally composed of actin filaments and myosin II motor proteins, together with other regulatory proteins such as formins, RhoA, and septins. These components form a dynamic, belt-like structure beneath the membrane at the equator of the dividing cell. The contractile ring produces the force that is needed for the ingression of the cleavage furrow. Myosin II proteins interact with actin filaments in the ring to generate this contractile force. This process is energized by hydrolysis of ATP. As the ring tightens, the plasma membrane is pinched inward, ultimately dividing the cytoplasm. The absence of the contractile ring would result in a failure of the cell to divide, leading to binucleated cells as well as other abnormalities.
Motor Proteins
Image source: Wikimedia Commons.
In a previous article, I described the role of motor proteins (kinesin and dynein) in the assembly and function of the mitotic spindle during eukaryotic cell division. I’d refer interested readers to that essay for a discussion of this astounding process. The absence of these motor proteins would severely compromise the transport and positioning of chromosomes, resulting in chromosomal misalignment during metaphase and difficulty in establishing a proper mitotic spindle. The consequence would be errors in chromosome segregation during anaphase.
Cdk and Cyclin Molecules
I have written previously about the role of cyclin-dependent kinases and cyclin molecules in cell cycle progression. I refer readers there for a review. The Cdk and cyclin molecules exhibit redundancy, meaning that they are not all individually necessary. For example, mouse knockouts of Cdk2, 3, 4, or 6 still retain viability. [7,8,9,10,11,12,13] Furthermore, yeast cells possess only a single Cdk, specifically Cdk1.14 Interestingly, double knockouts involving combinations of Cdk2 and 4, or Cdk4 and 6, result in embryonic lethality, though a double knockout of Cdk2 and 6 does not. [15,16] It appears, then, that the pair Cdk2 and 4 and the pair Cdk4 and 6 can substitute for one another.17 However, Cdk1 appears to be essential, and knocking it out arrests development at the blastocyst stage. [18]
Cdk molecules themselves are activated by the binding of cyclin molecules. Without those cyclins, the Cdks would be inactive, resulting in cell cycle arrest. Though there is redundancy here too (and thus not all cyclins are indispensable to successful division), the absence of cyclin B (which activates Cdk1 to drive progression into mitosis), would impair the transition from G2 to M phase. In other words, the cell could not enter mitosis. This is corroborated by experimental knockout studies of cyclin B in mouse embryos, leading to the arrest of the cell cycle in G2 after as little as two divisions. [19,20]
Checkpoints
I have written previously about the various cell cycle checkpoints — i.e., the G1 (restriction) checkpoint, G2 (DNA damage) checkpoint, and spindle assembly checkpoint (see my articles on these here, here and here). These are also essential for successful cell division. For instance, without the mitotic checkpoint complex, the cell’s ability to monitor spindle assembly would be abolished — drastically increasing the risk of cells proceeding through division with spindle defects, the result of which would be chromosome missegregation and aneuploidy. The absence of the G1 checkpoint would enable damaged DNA to enter S phase, which could lead to the propagation of mutations as well as genomic instability. The loss of the G2 checkpoint would allow cells with DNA damage to enter into mitosis, leading to the division of cells with unrepaired genetic material, as well as a greatly increased risk of chromosome aberrations. Without the DNA damage checkpoint in S phase, replication of damaged DNA would occur, resulting in the propagation of mutations and thus an elevated risk of genetic abnormalities in the daughter cells.
Irreducibly Complex
As seen from the cursory discussion above, various components of the mitotic cell division apparatus are indispensable for the system to work. This makes the eukaryotic cell division irreducibly complex, rendering it resistant to explanations in terms of blind, evolutionary processes. Any system that achieves a complex higher-level objective by means of various well-matched interacting components requires foresight to come about. In a subsequent article, I will discuss how the challenge to evolutionary accounts of the origins of eukaryotic cell division extends much deeper than this.
Notes
1. McLatchie J (2024) Phylogenetic Challenges to the Evolutionary Origin of the Eukaryotic Cell Cycle. BIO-Complexity 2024 (4):1–19 doi:10.5048/BIO-C.2024.4.
2. Kumada K, Yao R, Kawaguchi T, Karasawa M, Hoshikawa Y, et al (2006) The selective continued linkage of centromeres from mitosis to interphase in the absence of mammalian separase. J Cell Biol. 172(6): 835-46. doi:10.1083/jcb.200511126
3. Wirth KG, Wutz G, Kudo NR, Desdouets C, Zetterberg A, et al (2006) Separase: a universal trigger for sister chromatid disjunction but not chromosome cycle progression. J Cell Biol. 172(6): 847-60. doi:10.1083/jcb.200506119
4. Wang J, Yin MZ, Zhao KW, Ke F, Jin WJ, et al (2017) APC/C is essential for hematopoiesis and impaired in aplastic anemia. Oncotarget. 8(38): 63360-63369. doi:10.18632/oncotarget.18808
5. Lu LY, Wood JL, Ye L, Minter-Dykhouse K, Saunders TL, Yu X, Chen J (2008) Aurora A is essential for early embryonic development and tumor suppression. J Biol Chem. 283(46): 31785-90. doi:10.1074/jbc.M805880200
6. Yuba-Kubo A, Kubo A, Hata M, Tsukita S (2005) Gene knockout analysis of two gamma-tubulin isoforms in mice. Dev Biol.282(2): 361-73. doi:10.1016/j.ydbio.2005.03.031
7. Berthet C, Aleem E, Coppola V, Tessarollo L, Kaldis P (2003) Cdk2 knockout mice are viable. Curr Biol. 13: 1775–1785. doi:10.1016/j.cub.2003.09.024
8. Ortega S, et al. (2003) Cyclin-dependent kinase 2 is essential for meiosis but not for mitotic cell division in mice. Nat Genet.35: 25–31. doi:10.1038/ng1232
9. Ye X, Zhu C, Harper JW (2001) A premature-termination mutation in the Mus musculus cyclin-dependent kinase 3 gene. Proc Natl Acad Sci USA. 98: 1682–1686. doi:10.1073/pnas.98.4.1682
10. Rane SG, et al. (1999) Loss of Cdk4 expression causes insulin-deficient diabetes and Cdk4 activation results in β-islet cell hyperplasia. Nat Genet. 22: 44–52. doi:10.1038/8751
11. Tsutsui T, et al. (1999) Targeted disruption of CDK4 delays cell cycle entry with enhanced p27Kip1 activity. Mol Cell Biol. 19: 7011–7019. doi:10.1128/MCB.19.10.7011
12. Hu MG, et al. (2009) A requirement for cyclin-dependent kinase 6 in thymocyte development and tumorigenesis. Cancer Res. 69: 810–818. doi:10.1158/0008-5472.CAN-08-2473
13. Malumbres M, et al. (2004) Mammalian cells cycle without the D-type cyclin-dependent kinases Cdk4 and Cdk6. Cell. 118: 493–504. doi:10.1016/j.cell.2004.08.002
14. Enserink JM, Kolodner RD (2010) An overview of Cdk1-controlled targets and processes. Cell Div. 5: 11. doi:10.1186/1747-1028-5-11
15. Malumbres M, et al. (2004) Mammalian cells cycle without the D-type cyclin-dependent kinases Cdk4 and Cdk6. Cell. 118: 493–504. doi:10.1016/j.cell.2004.08.002
16. Berthet C, Kaldis P (2007) Cell-specific responses to loss of cyclin-dependent kinases. Oncogene 26: 4469–4477. doi:10.1038/sj.onc.1210243
17. Satyanarayana A, Kaldis P (2009) Mammalian cell-cycle regulation: Several Cdks, numerous cyclins and diverse compensatory mechanisms. Oncogene. 28:2925–2939. doi:doi.org/10.1038/onc.2009.170
18. Diril MK, Ratnacaram CK, Padmakumar VC, Du T, Wasser M, Coppola V, Tessarollo L, Kaldis P (2012) Cyclin-dependent kinase 1 (Cdk1) is essential for cell division and suppression of DNA re-replication but not for liver regeneration. Proc Natl Acad Sci U S A. 109(10): 3826-31. doi:10.1073/pnas.1115201109
19. Berthet C, et al. (2006) Combined loss of Cdk2 and Cdk4 results in embryonic lethality and Rb hypophosphorylation. Dev Cell. 10: 563–573. doi:10.1016/j.devcel.2006.03.004
20. Strauss B, Harrison A, Coelho PA, Yata K, Zernicka-Goetz M, Pines J (2018) Cyclin B1 is essential for mitosis in mouse embryos, and its nuclear export sets the time for mitosis. J Cell Biol. 217(1): 179-193. doi:10.1083/jcb.201612147
This article was originally published at Evolution News & Science Today, on November 27th, 2024.