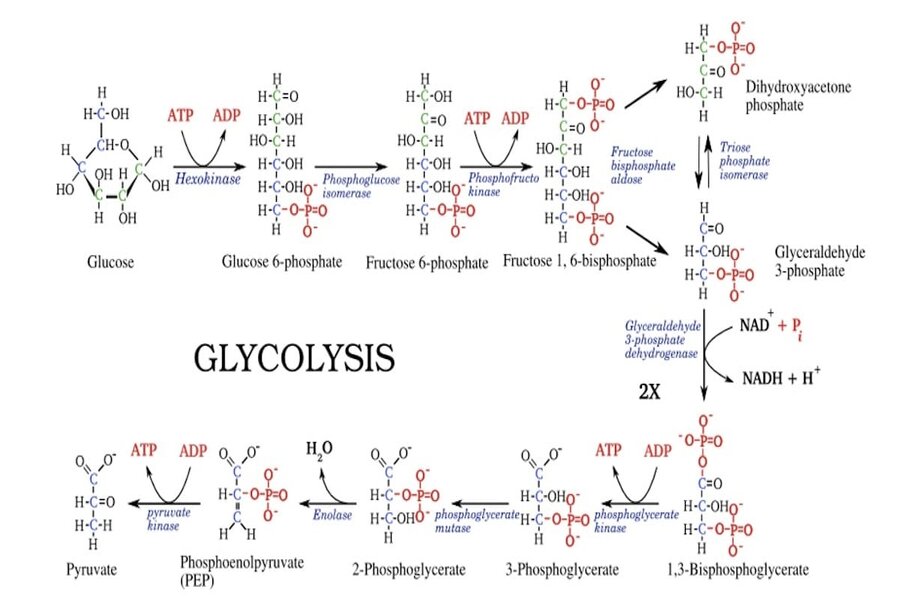
The purpose of cellular respiration is to convert the energy stored in glucose into adenosine triphosphate (ATP), the primary energy currency of the cell. Cellular respiration occurs in three main stages. Glycolysis involves the breakdown of glucose into pyruvate, producing a small amount of ATP. The citric acid cycle further breaks down pyruvate into carbon dioxide, generating NADH and FADH2. The final step of cellular respiration is the electron transport chain and oxidative phosphorylation, which produce a large amount of ATP, as well as water as a byproduct. In this article, I will discuss features of the glycolytic pathway that resist explanation by unguided evolutionary processes and which provide evidence of intelligent design.
The first step in cellular representation (glycolysis) is represented by the figure at the top. Glycolysis is ubiquitous across all living organisms. As shown in the figure, glycolysis involves the conversion of glucose, through a series of intermediates, to pyruvate. This pyruvate is then transported into the mitochondria where it is converted into acetyl-CoA by the enzyme pyruvate dehydrogenase. This process also produces NADH and releases one molecule of carbon dioxide (CO2). The acetyl-CoA then feeds into the citric acid cycle, where it is further oxidized, generating more NADH, FADH2, and ATP (or GTP).
Incremental Evolution?
Glycolysis has been proposed to be the first biochemical pathway to arise in evolution. Among the reasons for this are the fact that glycolysis is found ubiquitously across the tree of life (so may be inferred to have been present in the last universal common ancestor). Moreover, glycolysis is an anaerobic reaction sequence, and thus is consistent with the absence of oxygen in the primitive Earth environment.
There are, however, significant challenges to a proposed evolutionary origin of the glycolysis pathway. For example, the conversion of glucose to pyruvate involves as many as ten independent enzymes, typically 300 to 500 amino acids in length. It is extremely implausible that ten enzymes with complementary activities could have arisen at essentially the same time. But could the pathway have evolved incrementally, either forwards or backwards? It is generally rejected that glycolysis arose backwards (i.e., with pyruvate being initially available, then its precursor, etc.) since it was not the oxidized pyruvate, but rather sugar, that would have been present in the early Earth environment. Moreover, every intermediate between glycose and pyruvate is phosphorylated (i.e., has one or two of its hydroxyl groups replaced by phosphate). This involves a condensation reaction (where a water molecule is eliminated). Given the difficulties of this type of reaction, it is questionable whether the various intermediates could have emerged abiotically in high enough quantities to facilitate the origin of glycolysis.
The more popular view is that glycolysis evolved incrementally in the forwards direction. This hypothesis, of course, relies on the presumption that the intermediates could have served their own independent utility. However, since glycolysis is generally thought to have arisen extremely early — before additional utility of the intermediates could have arisen — it seems unlikely that the intermediates could have had independent usages.
Causal Circularity
Notice that the process of glycolysis consumes two ATP molecules — one at the glucose to glucose 6-phosphate step (catalyzed by hexokinase) and one at the fructose 6-phosphate to fructose 1,6-bisphosphate step (catalyzed by phosphofructokinase). The overall ATP yield of glycolysis is four (although many more ATPs will be produced later on), while two are consumed — making the net yield two ATPs. In order for ATP to be produced, ATP must first be consumed. This presents a causal circularity challenge to an evolutionary account of the origins of glycolysis. Strikingly, this causal circularity of ATP being required to manufacture more ATP appears to be ubiquitous across life. [1] How could the process of glycolysis be established without an initial supply of ATP? Moreover, after the consumption of the first ATP, there are at least five additional steps (each involving its own enzyme) before any further ATP is produced), and nine before there is a net yield of ATP. Given that natural selection lacks foresight, this renders it extremely implausible that the enzymes early on in the glycolytic pathway could have served any benefit in the absence of the enzymes later in the pathway.
Excluding Water
Of the ten enzymes involved in glycolysis, six catalyze reactions that involve a phosphate group transfer. For a phosphate to react with a hydroxyl group of water to form phosphoric acid is just as energetically favorable as for it to react with the hydroxyl or a sugar or ADP. But this would be of no evolutionary advantage. Thus, water must be excluded from the enzymes’ active sites to prevent hydrolytic reactions from occurring. This is achieved through a mechanism involving conformational changes that resemble a “hinge motion.” Initially, the enzyme’s active site assumes an open conformation, allowing the substrate to enter. When the substrate binds to the active site, it induces a conformational change, causing the enzyme to undergo a “closing” motion, with the domains of the enzyme coming together, effectively shielding the active site. This motion not only secures the substrate but also excludes water from entering the active site.
This phenomenon underscores the engineering sophistication — and the degree of amino acid specificity — of these enzymes. Since the exclusion of water is absolutely critical to the occurrence of the appropriate reactions, there would be no use in having a partly formed enzyme (i.e., one that could catalyze the phosphorylation reaction but failed to exclude water). This casts further doubt on the ability of incremental adaptations to account for the glycolytic pathway.
Relationship Between Enzymes?
A further issue is that, if indeed glycolysis were one of the earliest metabolic pathways to evolve, one might expect that at the time of its origin there existed only a small repertoire of enzymes. Moreover, the compounds on which these enzymes act have similar structures. This might lead us to predict that the enzymes involved in glycolysis are evolutionarily related to one another. However, as Keith Webster notes [2]:
Sequence and crystallographic data favor the divergent evolution of for example monophosphoglycerate mutase and diphosphoglycerate mutase, and possibly glyceraldehyde-3-P dehydrogenase and phosphoglycerate kinase from respective common ancestors, but convergence appears to have played a greater role in the development of all of the other 11 enzymes(Fothergill-Gilmore, 1986; Fothergill-Gilmore and Watson,1989). For example, there is no evidence of a common ancestor for any of the four glycolytic kinases or of the seven enzymes that bind nucleotides, with the exception of those mentioned above. Rather, it seems likely that the pathway resulted from the chance assembly of independently evolving enzymes and genes, probably in association with the co-evolution of other functions and linked pathways. [Emphasis added.]
This seems contrary to what might be predicted on an evolutionary account of the origins of glycolysis.
Intelligent Design
Multiple challenges confront an explanation of the glycolytic pathway in terms of unguided evolutionary mechanisms. The complexity and engineering sophistication comport much better with the hypothesis of design. In particular, the causal circularity of ATP being required to make more ATP is difficult to account for by a stepwise evolutionary process. On the other hand, this sort of phenomenon is totally unsurprising on the supposition of the involvement of an intelligent mind.
Notes
1. Kun A, Papp B, Szathmáry E. Computational identification of obligatorily autocatalytic replicators embedded in metabolic networks. Genome Biol. 2008;9(3): R51.
2. Webster KA. Evolution of the coordinate regulation of glycolytic enzyme genes by hypoxia. J Exp Biol. 2003 Sep; 206(Pt 17):2911-22.
This article was originally published at Evolution News & Science Today, on September 20, 2024.